Pendahuluan

Banyak yang
diketahui tentang persyaratan nutrisi kualitatif serangga. Aspek kuantitatif
nutrisi serangga kurang mendapat perhatian, dan ada beberapa studi tentang
tingkat asupan dan efisiensi pemanfaatan makanan. Secara khusus, relatif
sedikit yang diketahui tentang asupan, kecernaan dan efisiensi konversi diet
yang ditetapkan. Pekerjaan kuantitatif dengan diet buatan biasanya hanya
melibatkan pengukuran jumlah nutrisi tertentu yang diperlukan per unit diet
(House, 1959, 1962, 1965a). Ini mendefinisikan hubungan antara persyaratan
untuk nutrisi tertentu (Sang, 1956, 1959), tetapi tidak mengatakan apa pun
tentang asupan makanan, persyaratan absolut, atau efisiensi pemanfaatan
makanan. Sebagian besar pengukuran asupan dan pemanfaatan dilakukan dengan
serangga yang memakan makanan alami. Informasi yang tersedia di bidang ini
bermanfaat, tetapi beberapa di antaranya sulit untuk ditafsirkan karena
pengukuran belum distandarisasi, metode bervariasi dalam akurasi dan
kebingungan telah dihasilkan dari penerapan indeks pemanfaatan yang berasal
dari nutrisi vertebrata. Oleh karena itu, pada pembahasan ini telah dimasukkan
diskusi yang luas tentang poin-poin ini.
Lafon (1951)
mengkaji beberapa studi sebelumnya tentang serangga ulat sutera, Bombyx mori.
Sebagian besar dari banyak studi pemanfaatan makanan oleh serangga ini
dirangkum oleh Legay (1957, 1958) dan Yokoyama (1963). Saya telah menghilangkan
diskusi sebagian besar pekerjaan ini karena akan berulang dalam konteks ulasan
ini. Makalah terbaru tentang Bombyx yang tidak dikutip oleh pengulas di atas
adalah sebagai berikut: Hassanein dan El Shaarawy (1962a, b), Mukaiyama dan Ito
(1962a, b), Shyamala et al. (1956), Takeuchi dan Kosaka (1961). Pengukuran Trouvelot
(1867) tentang efisiensi makanan di Antheraea polyphemus adalah sejarah yang
menarik tetapi terlalu kasar untuk digunakan.
Serangga adalah
kelompok yang memakan berbagai zat organik yang sangat beragam. Pada saat yang
sama sebagian besar spesies menunjukkan tingkat spesifisitas yang tinggi dalam
pilihan makanan mereka. Gordon (1959) berhipotesis: "Persaingan dan
seleksi alam secara bertahap mendorong dan mengikat masing-masing spesies ke
pasokan makanan khusus yang dapat digunakan lebih efisien daripada
kompetitornya." Namun demikian, persyaratan nutrisi kualitatif dari
serangga yang tumbuh tampaknya relatif seragam ( Fraenkel, 1953, 1959; House,
1962). Tampaknya jelas bahwa perbedaan nutrisi adaptif harus dicari pada
tingkat kuantitatif dan bahwa nutrisi komparatif yang berarti dari serangga
tidak akan muncul sampai penelitian kuantitatif ditekankan. Penentuan
persyaratan absolut untuk konstituen makanan tergantung pada pengukuran asupan.
Perbedaan dalam efisiensi makanan hanya dapat ditunjukkan dengan mengukur
asupan dan pertumbuhan. Kecernaan juga harus diukur karena dapat diperkirakan
sangat bervariasi dengan makanan yang berbeda. Efisiensi dengan mana makanan
yang dicerna digunakan untuk pertumbuhan akan bervariasi tidak hanya dengan kebutuhan
pemeliharaan energi tetapi juga dengan keseimbangan nutrisi (Gordon, 1959).
Ukuran asupan
dan pemanfaatan telah memainkan peran kecil dalam studi klasik nutrisi serangga
meskipun kegunaannya dalam hubungan ini jelas. Sebagai contoh, contoh pertumbuhan
yang buruk mungkin bukan karena kekurangan nutrisi dari makanan tetapi karena
rendahnya asupan karena tidak adanya fagostimulan non-nutrisi. Sebaliknya,
penambahan nutrisi dengan aktivitas fagostimulatori dapat menyebabkan
peningkatan pertumbuhan meskipun nutrisi tersebut tidak diperlukan atau
digunakan (Dadd, 1960). Sang (1959) mengemukakan bahwa mungkin ada lebih dari
satu diet optimal untuk spesies tertentu. Ukuran asupan dan pemanfaatan dapat
memberikan indikasi akan hal ini, karena pola pemanfaatan mungkin berbeda
walaupun diet memiliki kemampuan yang sama untuk mendukung pertumbuhan.
Misalnya, asupan rendah mungkin diimbangi dengan kecernaan tinggi atau
pemanfaatan tinggi makanan yang dicerna untuk pertumbuhan atau sebaliknya.
Kecernaan yang buruk mungkin diimbangi dengan pemanfaatan makanan yang dicerna
secara efisien atau sebaliknya.
Salah satu
perhatian utama ekologi modern adalah penjelasan hubungan energetik di dalam
dan di antara masyarakat. Pengetahuan tentang efisiensi makanan serangga dengan
demikian sangat penting bagi ekologi karena, dalam hal jumlah atau biomassa,
serangga adalah komponen utama dari hampir semua komunitas air darat atau air
tawar. Engelmann (1966) meninjau bidang energetika terestrial dan membahas
signifikansi ekologisnya. Penulis juga telah membahas pentingnya data efisiensi
makanan dalam energi ekologis.
Indikasi Konsumsi, Pertumbuhan dan Pemanfaatan
Pemahaman
keseluruhan tentang pemanfaatan makanan membutuhkan jawaban atas
pertanyaan-pertanyaan berikut: Pada tingkat berapa makanan dimakan? Berapa
banyak dari makanan yang dimakan sebenarnya dicerna? Bagian mana dari makanan
yang dimasukkan sebagai zat tubuh? Pertanyaan-pertanyaan ini tidak dapat
dijawab tanpa metode yang baik dan akurat untuk menentukan berat makanan yang dicerna,
berat tinja yang sesuai dengan makanan yang dicerna dan berat yang diperoleh
serangga selama percobaan. Ketiga pengukuran ini mendasar dan perlu untuk
perhitungan laju pemberian makan, daya cerna dan efisiensi konversi makanan
menjadi zat tubuh. Hal ini berlaku apakah pertanyaannya adalah pemanfaatan
seluruh makanan (berat segar), bahan kering, energi, karbohidrat, nitrogen atau
konstituen gizi lain dari makanan.
Indeks yang
disebutkan di bawah ini dapat dihitung berdasarkan berat segar atau kering,
tetapi perlu dicatat bahwa indeks bobot segar dan kering tidak sebanding karena
persentase bahan kering makanan, tinja dan serangga cenderung berbeda.
Dalam karya klasiknya tentang faktor pertumbuhan aksesori Hopkins (1912) menunjukkan bahwa jumlah absolut tidak dapat digunakan untuk membandingkan asupan tikus yang tumbuh pada tingkat yang berbeda-bahwa perbandingan yang valid dapat dibuat hanya berdasarkan tingkat asupan relatif terhadap rata-rata. berat hewan selama periode menyusui. Indeks konsumsi (C.I.), oleh karena itu, dihitung sebagai:

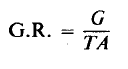




A. Konsumsi dan Pertumbuhan
1. Indeks konsumsi (Consumption index)Dalam karya klasiknya tentang faktor pertumbuhan aksesori Hopkins (1912) menunjukkan bahwa jumlah absolut tidak dapat digunakan untuk membandingkan asupan tikus yang tumbuh pada tingkat yang berbeda-bahwa perbandingan yang valid dapat dibuat hanya berdasarkan tingkat asupan relatif terhadap rata-rata. berat hewan selama periode menyusui. Indeks konsumsi (C.I.), oleh karena itu, dihitung sebagai:

Berat rata-rata hewan dihitung paling akurat dari area di bawah kurva pertumbuhannya yang ditentukan oleh integrasi atau pengukuran langsung. Rata-rata tertimbang dari bobot harian akan memberikan nilai yang hampir sama jika kurva pertumbuhannya halus (Waldbauer, 1964).
Indeks konsumsi lain telah diusulkan. Lafon (1951) menggunakan mg makanan yang dimakan per g serangga (berat akhir) dalam tiga puluh hari. Kegunaan indeks ini sulit untuk dilihat karena mengabaikan berat awal serangga. Misalnya, seekor serangga yang makan 500 mg saat tumbuh dari 10 hingga 100 mg jelas makan pada tingkat yang lebih besar relatif terhadap beratnya dibandingkan dengan seekor serangga yang makan dalam jumlah yang sama dalam waktu yang sama dengan pertumbuhan dari 50 hingga 100 mg. Legay (1957) mengusulkan mg makanan yang dimakan per g serangga (berat awal) per hari. Dalam hal ini dua serangga dengan berat awal yang sama yang memakan jumlah makanan yang sama pada waktu yang sama akan memiliki indeks yang identik bahkan jika satu tumbuh lebih dari yang lain. Smith (1959) menggunakan berat makanan yang dimakan dibagi dengan kenaikan berat badan. Ini menghasilkan indeks besar tak terhingga jika serangga makan tetapi tidak tumbuh, ekspresi dramatis dari kegagalan untuk tumbuh tetapi bukan ukuran yang berguna dari laju pemberian makan.
Lihat Bagian IV untuk diskusi lebih lanjut tentang tingkat pemberian makan.
2. Tingkat pertumbuhan (Growth Rate)
Tingkat pertumbuhan relatif (G.R.) dihitung sebagai:
Pekerja yang mengukur efisiensi pemanfaatan serangga biasanya tidak menghitung tingkat pertumbuhan sebenarnya (lihat, bagaimanapun, Waldbauer, 1964).
B. Perkiraan Kecernaan dan Efisiensi Konversi Makanan
1. Konversi makanan yang dicerna dihitung sebagai:
Efisiensi konversi makanan yang dicerna menjadi zat tubuh (E.C.I.)

E.C.I. adalah ukuran keseluruhan dari kemampuan serangga untuk memanfaatkan makanan yang ditelan untuk pertumbuhan. E.C.I. akan bervariasi dengan daya cerna suatu makanan dan jumlah proporsional dari porsi yang dapat dicerna dari makanan tersebut yang, di satu sisi, dikonversi menjadi bahan tubuh dan, di sisi lain, dimetabolisme menjadi energi untuk mempertahankan kehidupan. Demikianlah E.C.I. akan naik dan turun dengan A.D. (perkiraan kecernaan) dan E.C.D. (efisiensi konversi makanan yang dicerna menjadi zat tubuh).
2. Perkiraan Kecernaan
Perkiraan kecernaan (A.D.) dihitung sebagai:
Banyak penulis termasuk House (1965a), Trager (1953) dan Waldbauer (1964) menyebut ukuran ini sebagai "koefisien kecernaan". Namun, ini menyesatkan karena, seperti yang akan dibahas di bawah, perbedaan antara berat makanan yang dicerna dan berat kotoran tidak mewakili jumlah yang sebenarnya dicerna. Perlu dicatat bahwa perkiraan daya cerna seperti yang didefinisikan di sini untuk serangga berbeda dengan daya cerna yang tampak sebagaimana didefinisikan untuk mamalia.
Meskipun mamalia mengeluarkan urin dan feses secara terpisah, tinja mereka tidak, secara tegas, hanya terdiri dari makanan yang tidak tercerna juga tidak mengandung semua makanan yang tidak tercerna. Gas-gas yang diproduksi dalam pencernaan, terutama C02 dan CH4, hilang. Mamalia yang melakukan diet bebas nitrogen terus mengeluarkan nitrogen dalam tinja. Nitrogen fekal metabolik ini diduga berasal dari sekresi usus dan puing-puing sel usus. Jadi, pada mamalia, kecernaan yang sebenarnya tidak diperoleh dengan cara mengurangi berat tinja dari berat makanan yang dicerna. Nilai yang diperoleh disebut sebagai digestibility yang jelas dan dianggap sebagai perkiraan yang masuk akal dari digestibility yang sebenarnya (Kleiber, 1961; Mitchell, 1964; Tyler, 1964). Kotoran serangga tentu tidak bebas dari produk metabolisme tinja; membran peritrofik adalah contoh nyata.
Kehadiran urin dalam tinja semakin mempersulit pengukuran kecernaan pada serangga. Perbedaan antara berat makanan yang dicerna dan berat tinja sebenarnya mewakili makanan yang disimpan atau limbah metabolik yang kurang dimetabolisme yang dikeluarkan dalam urin atau sebagai produk metabolisme tinja. Dengan demikian, formula di atas menghasilkan apa yang disebut perkiraan kecernaan, nilai yang selalu lebih rendah dari kecernaan yang tampak. Besarnya perbedaan tergantung pada jumlah relatif urin dalam tinja. Burung mencampurkan urin dan feses di kloaka dan dengan demikian menimbulkan masalah yang sama (Kendeigh, 1949; Mitchell, 1964). Kendeigh mengacu pada energi dalam makanan yang dicerna kurang dari energi dalam kotoran sebagai energi yang dimetabolisme.
Dapat dikatakan bahwa penggunaan "kecernaan" - bahkan dalam arti "kecernaan yang diperkirakan" - tidak tepat kecuali satu yang mengandung paling tidak isi urin dari tinja. Namun, alternatif yang tersedia, "retensi" (Evans, 1939b) dan "pemanfaatan" (Kasting dan McGinnis, 1959; Hirano dan Ishii, 1962) juga tidak tepat. Selanjutnya, "pemanfaatan" harus dicadangkan sebagai istilah umum yang mencakup pencernaan, metabolisme dan konversi menjadi zat tubuh.
Perkiraan daya cerna yang jelas dapat diperoleh jika estimasi kadar urin dikurangi dari berat tinja. Saya akan merujuk pada nilai yang diperoleh sebagai koefisien kecernaan yang tidak jelas (C.A.D.).
Penentuan C.A.D. mungkin merupakan penyempurnaan yang tidak perlu dengan sebagian besar serangga yang memakan makanan dengan kandungan protein sedang. Kotoran mereka mungkin akan mengandung urin yang relatif sedikit, dan dengan demikian perbedaan antara A.D. dan C.A.D. kemungkinan sangat kecil. Komponen utama dari urin serangga terestrial adalah asam urat (lih. Patton, 1953; Wigglesworth, 1965). Analisis tinja serangga fitopag menunjukkan kadar asam urat yang rendah. Ini berkisar dari 0,51% untuk instar 1 turun ke 0,24% untuk instar ke-5 di tinja kering Bombyx mori (Hiratsuka, 1920). Pengurangan kandungan asam urat dari feses menghasilkan peningkatan A.D. yang dapat diabaikan. Ia dinaikkan dari 46,0% menjadi 46,4% untuk instar 1 dan dari 36,5% menjadi 36,7% untuk instar kelima (dihitung dari data Hiratsuka, 1920). Kotoran kering serangga pemakan daun lainnya juga rendah asam urat, 0,62% untuk instar ke-4 dan 0,52% untuk instar ke-5 dari Neodiprion sertifer (Janda, 1961), dan sekitar 4% untuk Melanoplus bivittatus (Brown, 1937a). Koreksi serupa untuk kandungan asam urat 4% dari feses larva Tenebrio molitor meningkatkan A.D. dari 46,3% menjadi 4,85% (Evans dan Goodliffe, 1939). Gupta dan Sinha (1960) menemukan konsentrasi asam urat yang lebih tinggi di dalam kotoran imago dari kumbang biji-bijian yang disimpan, sekitar 12% dari berat kering untuk Sitophilus granarius, 18% untuk Tribolium confusum dan hampir 20% untuk Cryptolestes ferrugineus.
Kotoran serangga yang memakan makanan tinggi protein cenderung mengandung persentase urin yang relatif lebih besar. Oleh karena itu, perkiraan kecernaan mungkin sangat berbeda dari koefisien kecernaan yang tampak. Larva ngengat pakaian yang berselaput, Tineola bisselliella, memakan rambut dan zat lain yang sebagian besar terdiri dari keratin. Kotoran keringnya mengandung sekitar 28% asam urat ditambah jumlah amonia dan urea yang jauh lebih kecil (Hollande dan Cordebard, 1926). Angka ini digunakan untuk memperbaiki data Titschack (1925) untuk Tineola (Tabel V) meningkatkan A.D. dari 29% menjadi C.A. sekitar 56%.
3. Konversi makanan yang dicerna
Efisiensi dengan makanan yang dicerna dikonversi menjadi zat tubuh (E.C.D.) dihitung sebagai:
E.C.D. akan menurun karena proporsi makanan yang dicerna yang dimetabolisme untuk energi meningkat. Demikianlah E.C.D. dipengaruhi oleh faktor-faktor yang mempengaruhi jumlah energi yang digunakan untuk pemeliharaan fungsi fisiologis atau dukungan aktivitas. E.C.D. tidak secara langsung tergantung pada kecernaan, tetapi itu bervariasi dengan tingkat asupan nutrisi. Karena kebutuhan pemeliharaan tetap kurang lebih konstan, proporsi makanan yang tersedia untuk pertumbuhan akan berkurang dengan berkurangnya asupan. E.C.D. juga bervariasi dengan nilai gizi makanan. Sebagai contoh, Hopkins (1912) menunjukkan bahwa kekurangan vitamin menyebabkan tikus membuang sejumlah besar makanan yang diserap (lihat Gordon (1959) tentang diskusi jelas tentang pentingnya efisiensi makanan.
Referensi : Waldbauer GP. 1968. The Consumption and Utilization of Food by Insects. Adv. In Insect Phys. 5(C):229–288. doi:10.1016/S0065-2806(08)60230-1.
Referensi : Waldbauer GP. 1968. The Consumption and Utilization of Food by Insects. Adv. In Insect Phys. 5(C):229–288. doi:10.1016/S0065-2806(08)60230-1.
Posting Komentar untuk "Konsumsi dan Pemanfaatan Makanan oleh Serangga : Bagian 1"