
Family yang Termasuk (20):
Agaonidae, Aphelinidae, Chalcididae, Elasmidae, Encyrtidae, Eucharitidae, Eulophidae, Eupelmidae, Eurytomidae, Leucospidae, Mymaridae, Ormyridae, Perilampidae, Pteromalidae, Rotoitidae, Signiphoridae, Tanaostigmatidae, Tetracampidae, Torymidae, Trichogrammatidae.Diagnosis:
Chalcidoid yang sepenuhnya bersayap dibedakan dari kebanyakan Hymenoptera lainnya dengan pengurangan venasi sayap depan. Paling banyak satu vena kompleks terjadi, terdiri dari vena submarginal, marginal, stigmal, dan postmarginal (Gambar 14, Bab 3). Kebanyakan chalcidoid juga memiliki sclerite yang terpisah, prepektus, yang memisahkan sebagian dari mesopleuron dari pronotum yang kurang lebih mirip pelana atau tapal kuda. Ini tidak seperti kebanyakan parasitoid Hymenoptera lainnya, yang tidak memiliki prepektus yang terbuka antara mesopleuron dan pronotum, dan memiliki pronotum sangat berkurang di medial sehingga seperti segitiga dalam tampilan lateral. Karena prepektus hadir antara pronotum dan mesopleuron di sebagian besar chalcidoid, pronotum biasanya tidak meluas ke tegula, tetapi betapa mencoloknya fitur ini tergantung pada ukuran prepektus. Posisi mesothoracic spiracle, jika terlihat, juga membedakan chalcidoid dari parasit Hymenoptera lainnya. Pada chalcidoid, mesothoracic spiracle berada di tepi dorsal pronotum, biasanya pada titik pronotum, prepektus, dan mesoscutum, tetapi setidaknya antara pronotum dan mesoscutum. Parasit Hymenoptera lainnya memiliki spiracle mesothoracic yang terletak di bawah tepi dorsal pronotum, baik antara pronotum dan mesopleuron (dalam beberapa taksa yang tersembunyi di bawah pronotallobe yang mencolok) atau pada pronotum itu sendiri dalam posisi relatif yang sama ini. Akibatnya, spiracle dan mesoscutum dipisahkan oleh sudut posterodorsal pronotum. Hampir semua chalcidoid juga memiliki struktur sensoris seperti membujur seperti punggungan (multiporous plate sensilla) pada satu atau lebih flagellomeres, dengan apeks sensilla yang diproyeksikan di atas permukaan, dan seringkali di luar puncak, dari flagellomere. Banyak chalcidoid juga memiliki kilau metalik, yang membedakannya dari sebagian besar microhymenoptera parasit lainnya. Boucek (1988a) memberikan tinjauan komprehensif struktur chalcidoid.

Keragaman biologis chalcidoid ditinjau secara rinci oleh Boucek (1988a), Gauld dan Bolton (1988), dan Bendel-Janssen (1977), dan sehingga hanya dibahas secara singkat untuk setiap famili. Kebanyakan chalcidoid adalah parasitoid atau, jarang, predator pada tahap yang belum matang atau, sangat jarang, pada dewasa dari 12 ordo Insekta, dua ordo Arachnida (Araneae dan Acari), dan satu keluarga Nematoda (Anguinidae). Ini mewakili jumlah pesanan yang sama yang disamaratakan oleh sisa Hymenoptera parasit bersama-sama. Beberapa chalcidoid adalah fitopag, baik sebagai pembentuk empedu atau pemakan biji, atau sebagai inquilines dalam galls spesies lain.
Anggota Chalcidoidea, atau Chalcidoidea plus Mymarommatidae, dapat ditunjukkan sebagai taksa monofiletik. Gibson (1986a) mendefinisikan Chalcidoidea atas dasar tiga atribut eksternal, diduga sinapomorfik. Karena anggota Mymarommatidae tidak memiliki tiga atribut ini, tetapi berbagi sinapomorfies putatif lainnya dengan chalcidoid, mereka dihipotesiskan sebagai sister grup Chalcidoidea sensu stricto. Gauld dan Bolton (1988) dan Naumann (1991) mengklasifikasikan mymarommatids sebagai sebuah famili di Chalcidoidea, sedangkan Delvare dan Aberlenc (1989) dan Noyes dan Valentine (1989b) mengklasifikasikan mereka sebagai superfamili yang terpisah dari Chalcidoidea. Dalam panduan ini, seekor mymarommatids; diklasifikasikan sebagai superfamili dan dikunci dalam kunci untuk superfamilies, tetapi mereka juga dikunci dalam kunci untuk keluarga Chalcidoidea karena ketidakstabilan saat ini dalam klasifikasi mereka.
Boucek (1988b) meninjau secara ekstensif sejarah klasifikasi chalcidoid. Sejak 1952 dari sembilan hingga 24 famili telah diakui dalam superfamili, dan saat ini peneliti belum mencapai konsensus untuk klasifikasi atau hubungan tingkat famili. Banyak famili tidak dapat didefinisikan oleh atribut unik atau bahkan kombinasi atribut jika fauna dunia diperlakukan. Selanjutnya, menggunakan kombinasi atribut untuk mengeluarkan taksa tingkat famili menghasilkan bentuk yang tampaknya tak berujung, meskipun sering langka, pengecualian atau menengah. Masalah-masalah ini telah lama diakui oleh para chalcidologists. Sebagaimana dinyatakan oleh Gordh (1979), klasifikasi didasarkan pada morfologi eksternal, dan chalcidoid sangat morfologis secara plastik. Hal ini mengakibatkan perbedaan pendapat atas batas-batas taksa yang lebih tinggi karena berbagai pekerja telah menafsirkan signifikansi karakter secara berbeda. Grissell (1980) juga mencatat bahwa karakter yang digunakan untuk membatasi taksa yang lebih tinggi sering bekerja dengan baik hanya untuk satu jenis kelamin dan umumnya tidak disjungtif, yaitu, mereka melakukan intergrade dan memotong dari waktu ke waktu di mana paling tidak diharapkan. Akibatnya, banyak keluarga chalcidoid mungkin mewakili taksa kenyamanan, berdasarkan kesamaan keseluruhan, lebih dari mereka mewakili silsilah evolusi monofiletik.
Klasifikasi famili yang digunakan di sini sebagian besar mengikuti Boucek (1988a), kecuali bahwa penggambaran Perilampidae dan Eucharitidae mengikuti Heraty dan Darling (1984). Karena banyak diagnosa famili tidak dapat dibuat secara ketat, atribut diagnostik dari unit terdefinisi paling inklusif dalam setiap famili (biasanya subfamili, jarang suku) diberikan, kecuali untuk Pteromalidae, yang umumnya dianggap sebagai dumping paraphyletic dari Chalcidoidea. Diagnosa famili disusun terutama oleh tingkat kesamaan struktural antara anggota, yang mungkin tidak mencerminkan hubungan filogenetik. Tidak ada upaya yang dilakukan untuk membuat diagnosa secara ketat dapat diperbandingkan karena ini akan cenderung mengaburkan atribut yang penting untuk pengakuan dari setiap takson tertentu. Namun, famili dengan anggota yang tampak serupa memiliki diagnosis yang lebih sebanding.
Regional keys to families (publications with an asterisk also include keys to genera) :
*Nikolskaya (1952) and *Medvedev (1978), European USSR; *Peck, Boucek and Hoffer (1964), Europe; Graham (1969), Europe-key to genera of Pteromalidae; Riek (1970), Australia; *Alayo and Hernandez (1978), Cuba; Prinsloo (1980), Ethiopian region; Yoshimoto (1984), Canada; *Subba Rao and Hayat (1985), Oriental region; *Boucek (1988a), Australasian region-keys to genera of 14 of 21 families; Gauld and Bolton (1988), Britain; Delvare and Aberlenc (1989), tropical Africa and America; *Noyes and Valentine (1989b), New Zealand-keys to genera of 12 of 16 families; Grissell and Schauff (1990), Nearctic region; Naumann (1991), Australia.

Komentar :
Chalcidoids ditemukan di semua daerah zoogeographic dan di semua habitat dari hutan khatulistiwa ke tundra paling utara, dari gurun ke kolam. Terlepas dari keberlimpahan mereka, mereka tetap menjadi salah satu kelompok parasit Hymenoptera termiskin, sebagian karena ukurannya yang kecil (kebanyakan panjangnya 3-5 mm atau kurang), keragaman morfologi dan biologis, dan kelimpahan numerik. Nominal (jumlahnya) sekitar 3300 genera dan 22 500 spesies yang telah dideskripsikan, di mana sekitar 2000 genera dan 18.500 spesies dianggap valid (Noyes 1990a). Jumlah spesies tentu hanya mewakili sebagian kecil dari keragaman sejati Chalcidoidea, dan perkiraan spesies chalcidoid 60000-100000 tampaknya tidak masuk akal (Noyes 1978, Gordh 1979). Perkiraan jumlah genus dan spesies yang diberikan untuk setiap keluarga dalam panduan ini didasarkan pada Noyes (1990a).Keragaman biologis chalcidoid ditinjau secara rinci oleh Boucek (1988a), Gauld dan Bolton (1988), dan Bendel-Janssen (1977), dan sehingga hanya dibahas secara singkat untuk setiap famili. Kebanyakan chalcidoid adalah parasitoid atau, jarang, predator pada tahap yang belum matang atau, sangat jarang, pada dewasa dari 12 ordo Insekta, dua ordo Arachnida (Araneae dan Acari), dan satu keluarga Nematoda (Anguinidae). Ini mewakili jumlah pesanan yang sama yang disamaratakan oleh sisa Hymenoptera parasit bersama-sama. Beberapa chalcidoid adalah fitopag, baik sebagai pembentuk empedu atau pemakan biji, atau sebagai inquilines dalam galls spesies lain.
Anggota Chalcidoidea, atau Chalcidoidea plus Mymarommatidae, dapat ditunjukkan sebagai taksa monofiletik. Gibson (1986a) mendefinisikan Chalcidoidea atas dasar tiga atribut eksternal, diduga sinapomorfik. Karena anggota Mymarommatidae tidak memiliki tiga atribut ini, tetapi berbagi sinapomorfies putatif lainnya dengan chalcidoid, mereka dihipotesiskan sebagai sister grup Chalcidoidea sensu stricto. Gauld dan Bolton (1988) dan Naumann (1991) mengklasifikasikan mymarommatids sebagai sebuah famili di Chalcidoidea, sedangkan Delvare dan Aberlenc (1989) dan Noyes dan Valentine (1989b) mengklasifikasikan mereka sebagai superfamili yang terpisah dari Chalcidoidea. Dalam panduan ini, seekor mymarommatids; diklasifikasikan sebagai superfamili dan dikunci dalam kunci untuk superfamilies, tetapi mereka juga dikunci dalam kunci untuk keluarga Chalcidoidea karena ketidakstabilan saat ini dalam klasifikasi mereka.
Boucek (1988b) meninjau secara ekstensif sejarah klasifikasi chalcidoid. Sejak 1952 dari sembilan hingga 24 famili telah diakui dalam superfamili, dan saat ini peneliti belum mencapai konsensus untuk klasifikasi atau hubungan tingkat famili. Banyak famili tidak dapat didefinisikan oleh atribut unik atau bahkan kombinasi atribut jika fauna dunia diperlakukan. Selanjutnya, menggunakan kombinasi atribut untuk mengeluarkan taksa tingkat famili menghasilkan bentuk yang tampaknya tak berujung, meskipun sering langka, pengecualian atau menengah. Masalah-masalah ini telah lama diakui oleh para chalcidologists. Sebagaimana dinyatakan oleh Gordh (1979), klasifikasi didasarkan pada morfologi eksternal, dan chalcidoid sangat morfologis secara plastik. Hal ini mengakibatkan perbedaan pendapat atas batas-batas taksa yang lebih tinggi karena berbagai pekerja telah menafsirkan signifikansi karakter secara berbeda. Grissell (1980) juga mencatat bahwa karakter yang digunakan untuk membatasi taksa yang lebih tinggi sering bekerja dengan baik hanya untuk satu jenis kelamin dan umumnya tidak disjungtif, yaitu, mereka melakukan intergrade dan memotong dari waktu ke waktu di mana paling tidak diharapkan. Akibatnya, banyak keluarga chalcidoid mungkin mewakili taksa kenyamanan, berdasarkan kesamaan keseluruhan, lebih dari mereka mewakili silsilah evolusi monofiletik.
Klasifikasi famili yang digunakan di sini sebagian besar mengikuti Boucek (1988a), kecuali bahwa penggambaran Perilampidae dan Eucharitidae mengikuti Heraty dan Darling (1984). Karena banyak diagnosa famili tidak dapat dibuat secara ketat, atribut diagnostik dari unit terdefinisi paling inklusif dalam setiap famili (biasanya subfamili, jarang suku) diberikan, kecuali untuk Pteromalidae, yang umumnya dianggap sebagai dumping paraphyletic dari Chalcidoidea. Diagnosa famili disusun terutama oleh tingkat kesamaan struktural antara anggota, yang mungkin tidak mencerminkan hubungan filogenetik. Tidak ada upaya yang dilakukan untuk membuat diagnosa secara ketat dapat diperbandingkan karena ini akan cenderung mengaburkan atribut yang penting untuk pengakuan dari setiap takson tertentu. Namun, famili dengan anggota yang tampak serupa memiliki diagnosis yang lebih sebanding.
Regional keys to families (publications with an asterisk also include keys to genera) :
*Nikolskaya (1952) and *Medvedev (1978), European USSR; *Peck, Boucek and Hoffer (1964), Europe; Graham (1969), Europe-key to genera of Pteromalidae; Riek (1970), Australia; *Alayo and Hernandez (1978), Cuba; Prinsloo (1980), Ethiopian region; Yoshimoto (1984), Canada; *Subba Rao and Hayat (1985), Oriental region; *Boucek (1988a), Australasian region-keys to genera of 14 of 21 families; Gauld and Bolton (1988), Britain; Delvare and Aberlenc (1989), tropical Africa and America; *Noyes and Valentine (1989b), New Zealand-keys to genera of 12 of 16 families; Grissell and Schauff (1990), Nearctic region; Naumann (1991), Australia.
Penggunaan kunci:
Kunci berupaya untuk memungkinkan identifikasi chalcidoids, atau setidaknya betina chalcidoids, untuk famili di dunia. Ini barangkali merupakan tugas Sisyphean karena keragaman morfologi besar yang ditemukan di banyak famili bahkan ketika ditinjau secara regional, apalagi basis dunia, dan penempatan famili tak tentu dari beberapa taksa. Untuk alasan-alasan ini anggota dari beberapa subfamilies, seperti yang saat ini ditujukan ke Agaonidae, dibedakan dalam kunci famili. Masalah keragaman morfologi juga menghasilkan kunci yang penuh dengan kata-kata "biasanya" dan "sering," dan konjungsi seperti "dan," "atau," "tetapi." Kata-kata yang memenuhi syarat ini penting untuk identifikasi spesimen yang benar dan dalam bentuk cetak tebal. Catatan kaki digunakan untuk menunjukkan pengecualian atau atribut yang tidak berlaku untuk taksa langka, yang harus dikunci melalui kuplet masing-masing. Informasi tambahan diberikan mengikuti kuplet untuk membantu mengenali masalah taksa. Sebagai contoh, beberapa anggota Pleurotroppopsis (Eulophidae) memiliki metafemur bergigi di bagian perut, tetapi mereka memiliki empat tarsomer dan dengan demikian akan dikunci melalui "aa" di dalam bait 1. Karena upaya kunci 'untuk mencakup keragaman dunia itu tidak mengabaikan pengecualian untuk lebih mudah. konstruksi, dan memang rumit. Pengguna kunci juga harus mencatat bahwa karena chalcidoid umumnya cukup kecil dan rapuh, keberhasilan dalam memasukkan spesimen tergantung pada seberapa baik diawetkan dan disiapkan spesimen, dan apakah perbesaran yang memadai dan pencahayaan yang sesuai digunakan. (Noyes (1990b) memberikan tinjauan yang sangat baik dan komprehensif teknik untuk membunuh, melestarikan, dan pemasangan chalcidoid untuk studi taksonomi.) Jika memungkinkan, pendeskripsi yang digunakan dalam kunci telah disederhanakan untuk menghindari jargon khusus; istilah sinonim yang digunakan dalam literatur chalcidoid dan pernyataan penjelasan tambahan untuk parset diberikan dalam tanda kurung.Ciri-ciri Superfamily Chalcidoidea
Bagian-bagian tubuh








Ciri-ciri Sesuai Kunci Taksonomi

a. Sayap depan tanpa sel tertutup dan hanya 1 atau 2 vena.
b. Badan sangat kecil (kurang dari 3 mm)
(beberapa CHALCIDOIDEA) |

a. Kepala dengan tanda berbentuk H gelap antara mata, ocelli,, dan
toruli.
b. Toruli biasanya lebih dekat dengan mata daripada satu sama lain
(masing-masing torulus biasanya kurang dari diameternya sendiri dari mata).
c. Sayap belakang mengintai secara mendasar (selaput sayap belakang tidak
meluas ke pangkal sayap) dan sayap kedepan tampaknya hanya dengan 1 vena.
(Tubuh biasanya di bawah 1,5 mm panjang, sebagian besar spesies jauh lebih
kecil). . . . . . . . . . . (MYMARIDAE) CHALCIDOIDEA
|
 
a. Pronotum (nol) dalam tampilan lateral biasanya dipisahkan dari
tegula (tg).
b. Mesothoracic spiracle (sp) (tidak selalu terlihat) terletak antara
pronotum dan margin lateral mesoscutum (ms), jauh dari margin anterior
mesopleuron (plz).
c. Tubuh sering dengan warna metalik, atau antena dengan setidaknya 1
menit, seperti cincin flagellomere hanya setelah pedicel, atau keduanya
(tubuh spesies yang lebih kecil sering lemah sclerotized, sehingga spesimen
kering udara sering runtuh di bagian kepala, mesosoma dan metasoma) . . . . .
. .. mayoritas CHALCIDOIDEA
|
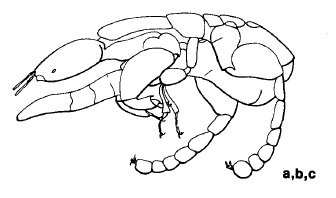
a. Jantan ; ditemukan secara eksklusif di dalam buah ara (buah
Ficus).
b. Antena lebih pendek dari kepala.
c. Metasoma sering panjang, lemah sclerotized, dan pucat.. . . . . . (AGAONIDAE) CHALCIDOIDEA
|

a. Pronotum (nol) dalam pandangan lateral tidak meluas ke tegula
(tg).
b. Integument biasanya tipis, terutama bagian dorsal dari metasoma;
tubuh hampir selalu roboh dalam spesimen kering-udara CHALCIDOIDEA
|
Sumber :
Gibson GAP. 1993. Superfamilies Mymarommatoidea and Chalcidoidea. Di dalam Goulet H, Huber JT, editor. Hymenoptera of the World: An identification guide to families. Ottawa: Canada Communication Group Publishing.
Grissell EE, Schauff ME. 1990. A Handbook of The Families of Nearctic Chalcidoidea (Hymenoptera). Washingthon DC: The Entomologycal Society of Washingthon.
Gibson GAP. 1993. Superfamilies Mymarommatoidea and Chalcidoidea. Di dalam Goulet H, Huber JT, editor. Hymenoptera of the World: An identification guide to families. Ottawa: Canada Communication Group Publishing.
Grissell EE, Schauff ME. 1990. A Handbook of The Families of Nearctic Chalcidoidea (Hymenoptera). Washingthon DC: The Entomologycal Society of Washingthon.
Posting Komentar untuk "Superfamily CHALCIDOIDEA"